Описание слайда:
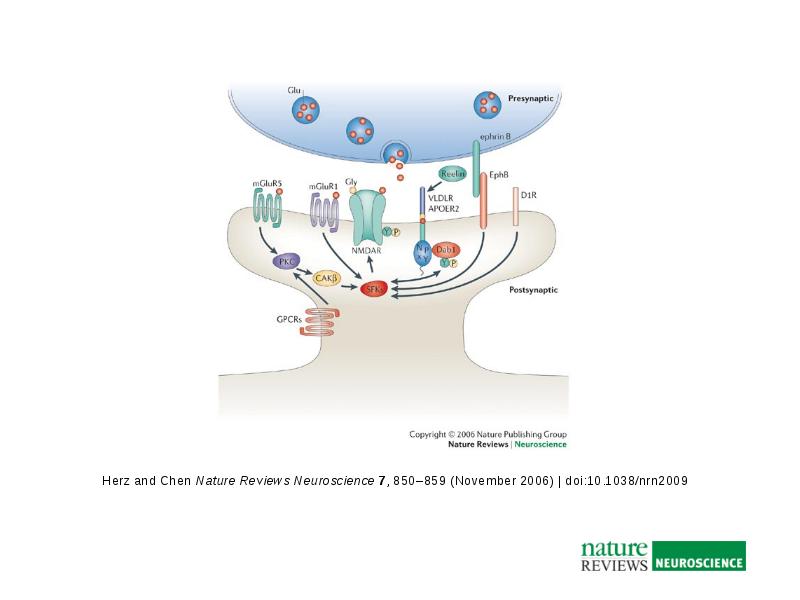
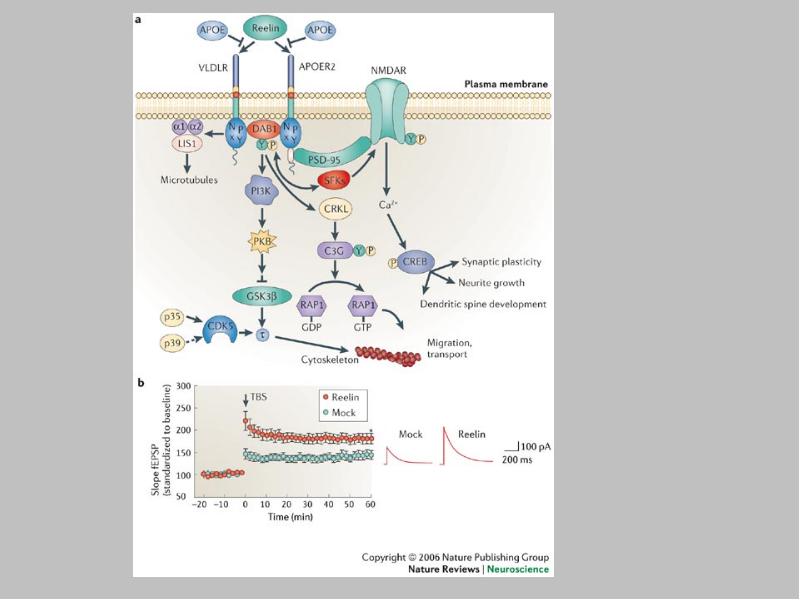
a | Reelin binds to lipoprotein receptors, the VLDLR and the APOER2, with high affinity at the cell surface. Binding of reelin to the receptors induces feed-forward activation of DAB1, an adaptor protein that interacts with NPxY motifs in both receptor tails. The clustering of DAB1 activates SRC family tyrosine kinases (SFKs), which potentiates tyrosine phosphorylation of DAB1 (Refs 32, 33). Phosphorylated DAB1 further activates phosphatidylinositol-3-kinase (PI3K) and subsequently protein kinase B (PKB)121. PKB activation inhibits the activity of glycogen synthase kinase 3 (GSK3). As a result, phosphorylation of is reduced, promoting microtubule stability. Tyrosine-phosphorylated (YP) DAB1 also recruits CRK-like (CRKL), which induces phosphorylation of a guanine nucleotide exchange factor, C3G122. Activated C3G promotes the formation of RAP1-GTP, which controls actin cytoskeleton rearrangement. Lissencephaly 1 (LIS1) is another binding partner of tyrosine-phosphorylated DAB1. It associates with a-subunits to form a Pafah1b complex, which regulates microtubule dynamics35, 123, 124, 125, 126, 127. Cyclin-dependent kinase 5 (CDK5) acts in parallel with reelin on numerous substrates, including microtubules. p35 and p39 are activating subunits of CDK5. APOER2 associates with postsynaptic density protein 95 (PSD-95), an abundant scaffolding protein in the PSD, through an alternatively spliced exon. This interaction is crucial for the coupling of the reelin signalling complex to the NMDA (N-methyl-D-aspartate) receptor (NMDAR)84, 86. Reelin-activated SFKs tyrosine phosphorylate the NMDAR on NR2 subunits, resulting in the potentiation of NMDAR-mediated Ca2+ influx. Elevated intracellular Ca2+ can activate the transcription factor cyclic AMP-response element binding protein (CREB), thereby potentially initiating the expression of genes that are important for synaptic plasticity, neurite growth and dendritic spine development86. b | Reelin treatment potentiates long-term potentiation in wild-type hippocampal slices82. Reelin treatment also enhances NMDAR-mediated whole cell current in wild-type CA1 pyramidal neurons86. Traces were recorded at a holding potential of +40 mV. fESP, field excitatory postsynaptic potential; TBS, theta burst stimulation. Panel a modified, with permission from Ref. 84 © (2005) Elsevier Science. Panels a and b (right) modified, with permission, from Ref. 82 © (2002) American Society for Biochemistry and Molecular Biology. Panel b (left) modified from Ref. 86 © (2005) Society for Neuroscience.
a | Reelin binds to lipoprotein receptors, the VLDLR and the APOER2, with high affinity at the cell surface. Binding of reelin to the receptors induces feed-forward activation of DAB1, an adaptor protein that interacts with NPxY motifs in both receptor tails. The clustering of DAB1 activates SRC family tyrosine kinases (SFKs), which potentiates tyrosine phosphorylation of DAB1 (Refs 32, 33). Phosphorylated DAB1 further activates phosphatidylinositol-3-kinase (PI3K) and subsequently protein kinase B (PKB)121. PKB activation inhibits the activity of glycogen synthase kinase 3 (GSK3). As a result, phosphorylation of is reduced, promoting microtubule stability. Tyrosine-phosphorylated (YP) DAB1 also recruits CRK-like (CRKL), which induces phosphorylation of a guanine nucleotide exchange factor, C3G122. Activated C3G promotes the formation of RAP1-GTP, which controls actin cytoskeleton rearrangement. Lissencephaly 1 (LIS1) is another binding partner of tyrosine-phosphorylated DAB1. It associates with a-subunits to form a Pafah1b complex, which regulates microtubule dynamics35, 123, 124, 125, 126, 127. Cyclin-dependent kinase 5 (CDK5) acts in parallel with reelin on numerous substrates, including microtubules. p35 and p39 are activating subunits of CDK5. APOER2 associates with postsynaptic density protein 95 (PSD-95), an abundant scaffolding protein in the PSD, through an alternatively spliced exon. This interaction is crucial for the coupling of the reelin signalling complex to the NMDA (N-methyl-D-aspartate) receptor (NMDAR)84, 86. Reelin-activated SFKs tyrosine phosphorylate the NMDAR on NR2 subunits, resulting in the potentiation of NMDAR-mediated Ca2+ influx. Elevated intracellular Ca2+ can activate the transcription factor cyclic AMP-response element binding protein (CREB), thereby potentially initiating the expression of genes that are important for synaptic plasticity, neurite growth and dendritic spine development86. b | Reelin treatment potentiates long-term potentiation in wild-type hippocampal slices82. Reelin treatment also enhances NMDAR-mediated whole cell current in wild-type CA1 pyramidal neurons86. Traces were recorded at a holding potential of +40 mV. fESP, field excitatory postsynaptic potential; TBS, theta burst stimulation. Panel a modified, with permission from Ref. 84 © (2005) Elsevier Science. Panels a and b (right) modified, with permission, from Ref. 82 © (2002) American Society for Biochemistry and Molecular Biology. Panel b (left) modified from Ref. 86 © (2005) Society for Neuroscience.