Описание слайда:
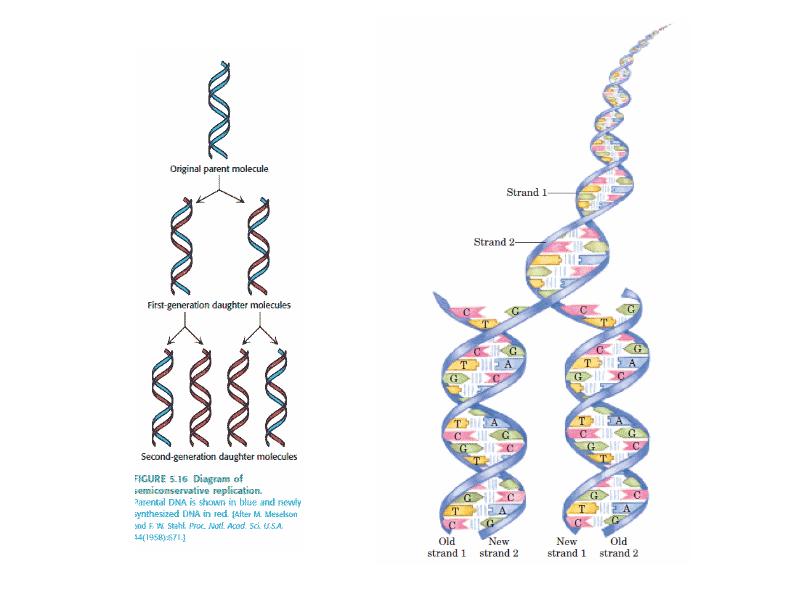
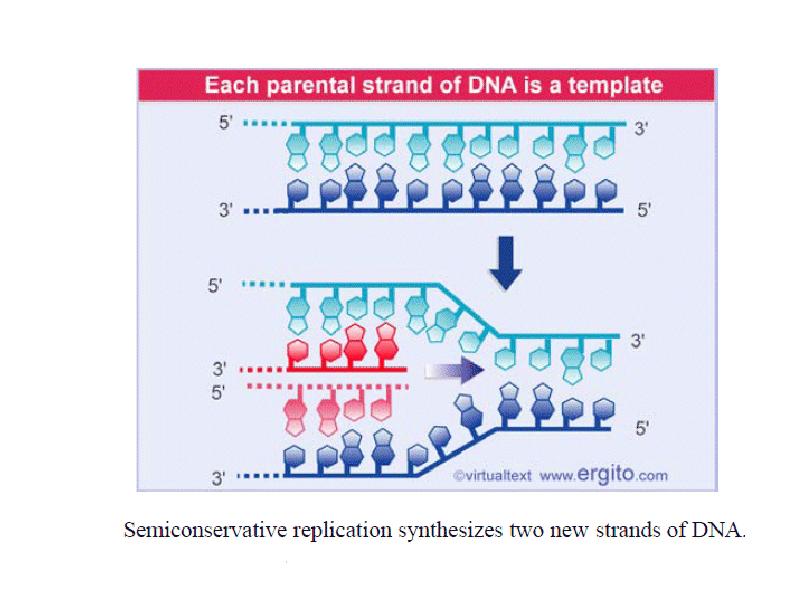
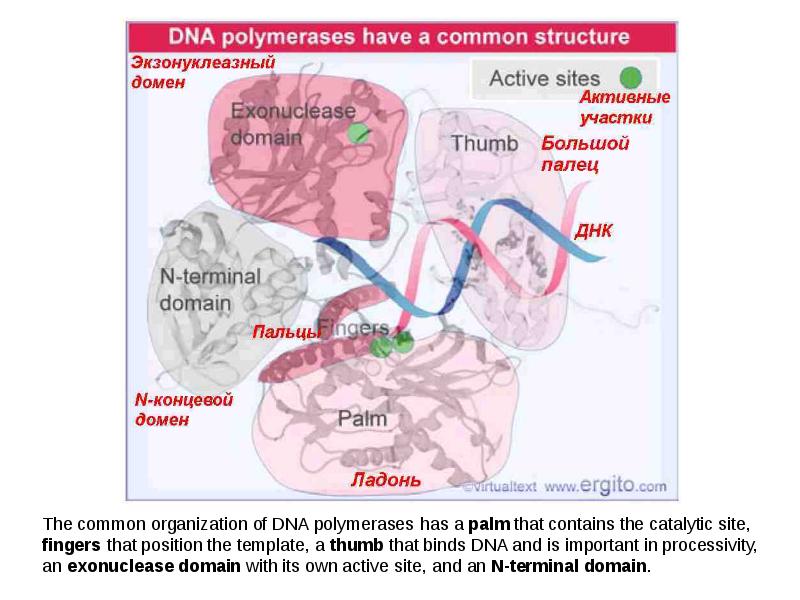
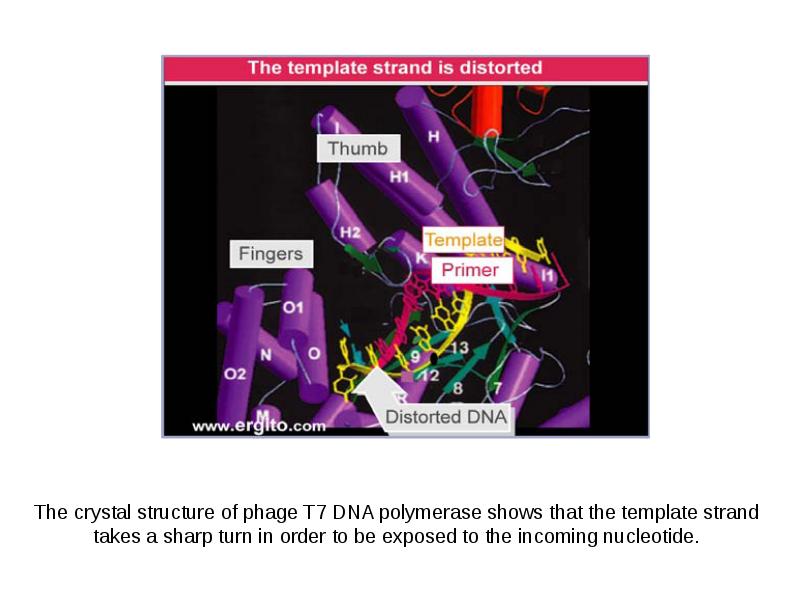
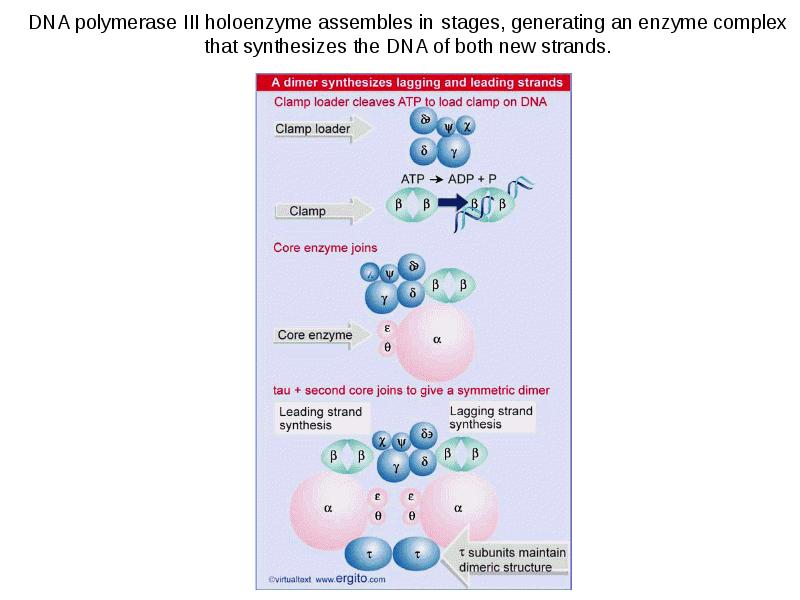
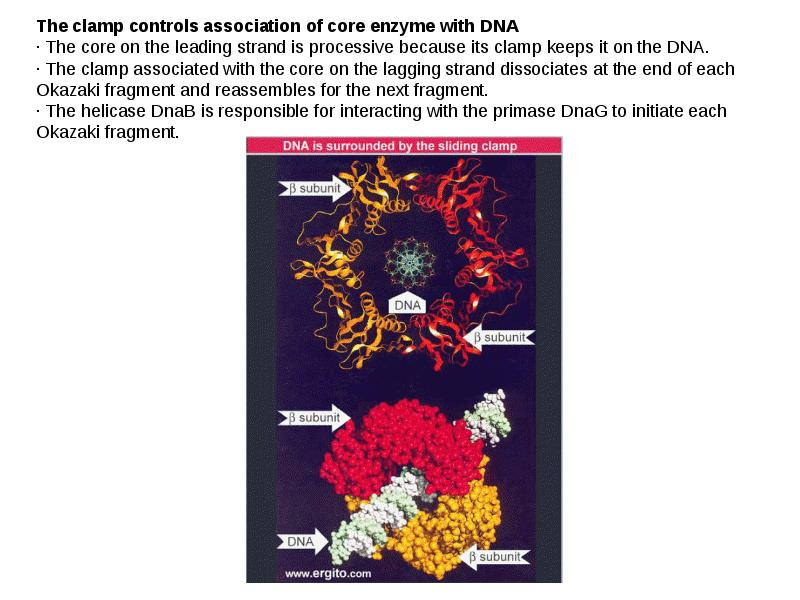
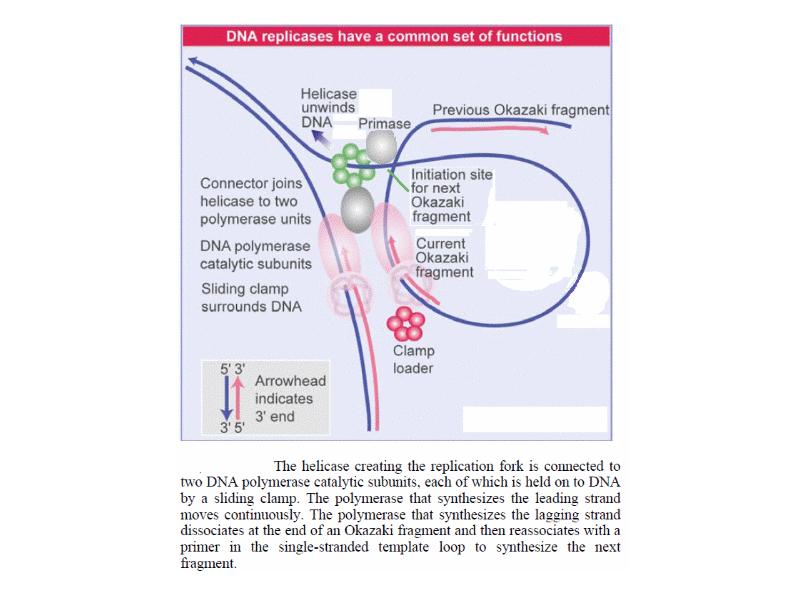
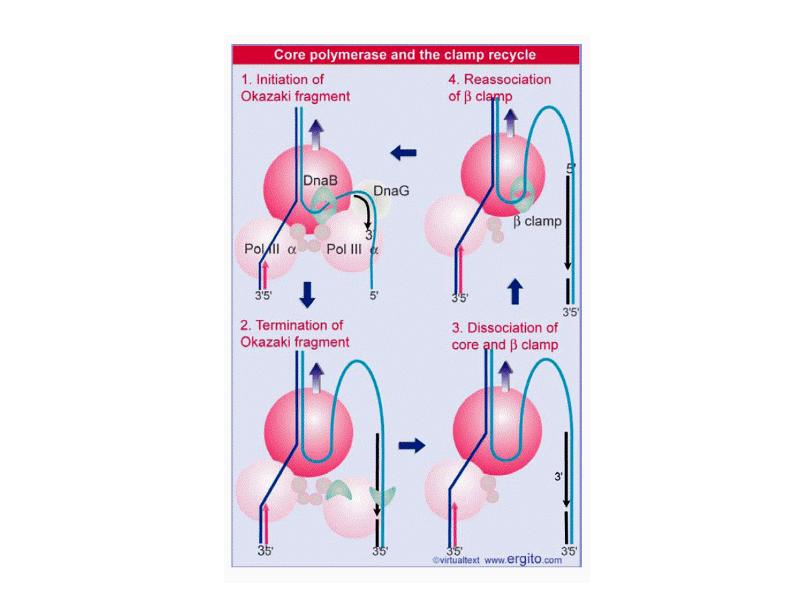
DNA REPLICATION Summary DNA synthesis occurs by semidiscontinuous replication, in which the leading strand of DNA growing 5 ¢ –3 ¢ is extended continuously, but the lagging strand that grows overall in the opposite 3 ¢ –5 ¢ direction is made as short Okazaki fragments, each synthesized 5 ¢ –3 ¢ . The leading strand and each Okazaki fragment of the lagging strand initiate with an RNA primer that is extended by DNA polymerase. Bacteria and eukaryotes each possess more than one DNA polymerase activity. DNA polymerase III synthesizes both lagging and leading strands in E. coli. Many proteins are required for DNA polymerase III action and several constitute part of the replisome within which it functions. The replisome contains an asymmetric dimer of DNA polymerase III; each new DNA strand is synthesized by a different core complex containing a catalytic ( a) subunit. Processivity of the core complex is maintained by the b clamp, which forms a ring round DNA. The clamp is loaded on to DNA by the clamp loader complex. Clamp/clamp loader pairs with similar structural features are widely found in both prokaryotic and eukaryotic replication systems. The looping model for the replication fork proposes that, as one half of the dimer advances to synthesize the leading strand, the other half of the dimer pulls DNA through as a single loop that provides the template for the lagging strand. The transition from completion of one Okazaki fragment to the start of the next requires the lagging strand catalytic subunit to dissociate from DNA and then to reattach to a b clamp at the priming site for the next Okazaki fragment. DnaB provides the helicase activity at a replication fork; this depends on ATP cleavage. DnaB may function by itself in oriC replicons to provide primosome activity by interacting periodically with DnaG, which provides the primase that synthesizes RNA. Phage T4 codes for a replication apparatus consisting of 7 proteins: DNA polymerase, helicase, single-strand binding protein, priming activities, and accessory proteins. Similar functions are required in other replication systems, including a HeLa cell system that replicates SV40 DNA. Different enzymes, DNA polymerase a and DNA polymerase d, initiate and elongate the new strands of DNA. The fX priming event also requires DnaB, DnaC, and DnaT. PriA is the component that defines the primosome assembly site (pas) for fX replicons; it displaces SSB from DNA in an action that involves cleavage of ATP. PriB and PriC are additional components of the primosome. The importance of the primosome for the bacterial cell is that it is used to restart replication at forks that stall when they encounter damaged DNA. The common mode of origin activation involves an initial limited melting of the double helix, followed by more general unwinding to create single strands. Several proteins act sequentially at the E. coli origin. Replication is initiated at oriC in E. Coli when DnaA binds to a series of 9 bp repeats. This is followed by binding to a series of 13 bp repeats, where it uses hydrolysis of ATP to generate the energy to separate the DNA strands. The pre-priming complex of DnaC-DnaB displaces DnaA. DnaC is released in a reaction that depends on ATP hydrolysis; DnaB is joined by the replicase enzyme, and replication is initiated by two forks that set out in opposite directions. Similar events occur at the lambda origin, where phage proteins O and P are the counterparts of bacterial proteins DnaA and DnaC, respectively. In SV40 replication, several of these activities are combined in the functions of T antigen. The availability of DnaA at the origin is an important component of the system that determines when replication cycles should initiate. Following initiation of replication, DnaA hydrolyzes its ATP under the stimulus of the b sliding clamp, generating an inactive form of the protein. Also, oriC must compete with the dat site for binding DnaA. Several sites that are methylated by the Dam methylase are present in the E. Coli origin, including those of the 13-mer binding sites for DnaA. The origin remains hemimethylated and is in a sequestered state for ~10 minutes following initiation of a replication cycle. During this period it is associated with the membrane, and reinitiation of replication is repressed. The protein SeqA is involved in sequestration and may interact with DnaA. After cell division, nuclei of eukaryotic cells have a licensing factor that is needed to initiate replication. Its destruction after initiation of replication prevents further replication cycles from occurring in yeast. Licensing factor cannot be imported into the nucleus from the cytoplasm, and can be replaced only when the nuclear membrane breaks down during mitosis. The origin is recognized by the ORC proteins, which in yeast remain bound throughout the cell cycle. The protein Cdc6 is available only at S phase. In yeast it is synthesized during S phase and rapidly degraded. In animal cells it is synthesized continuously, but is exported from the nucleus during S phase. The presence of Cdc6 allows the MCM proteins to bind to the origin. The MCM proteins are required for initiation. The action of Cdc6 and the MCM proteins provides the licensing function.