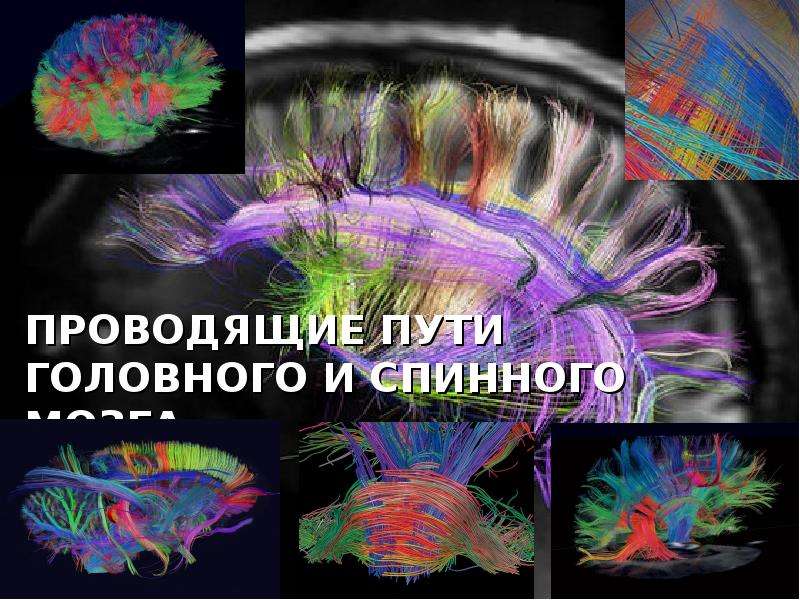
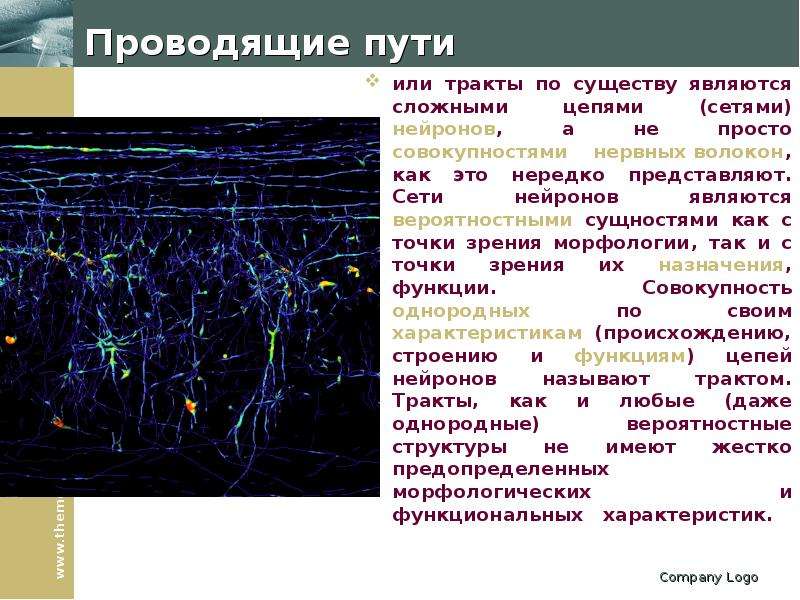
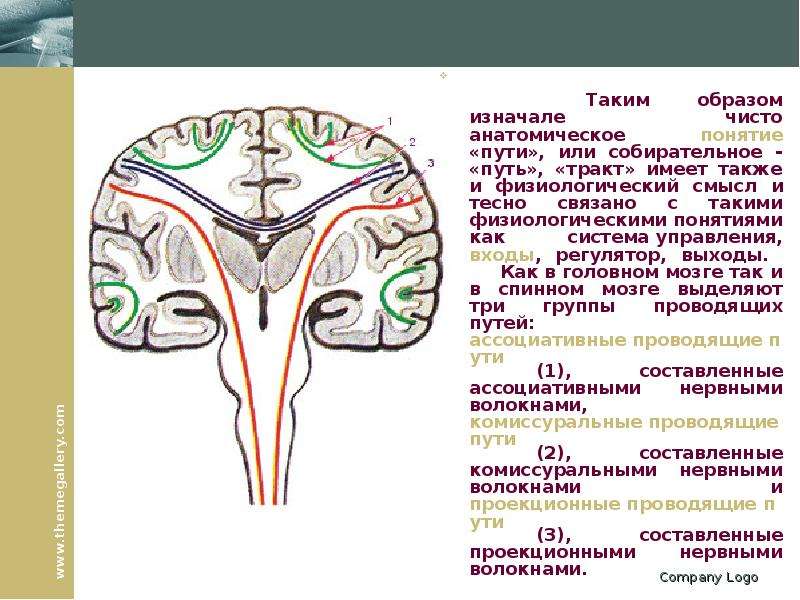
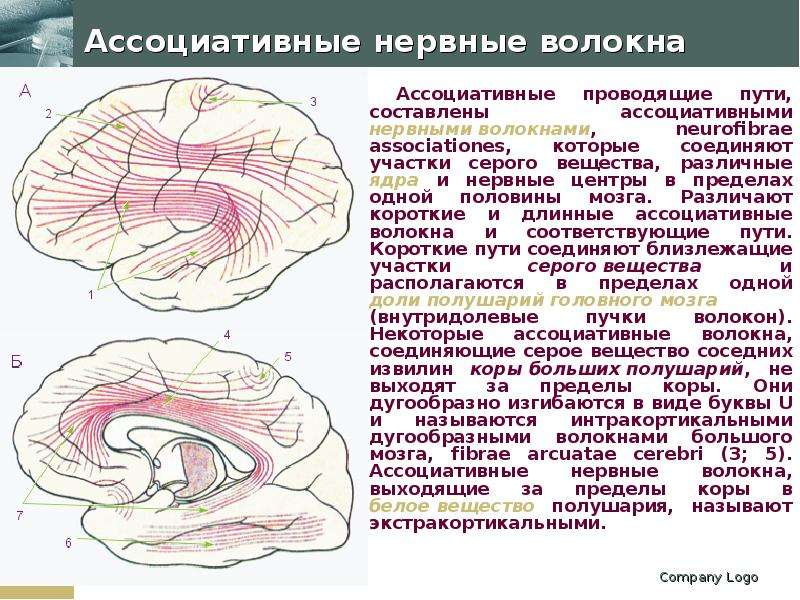
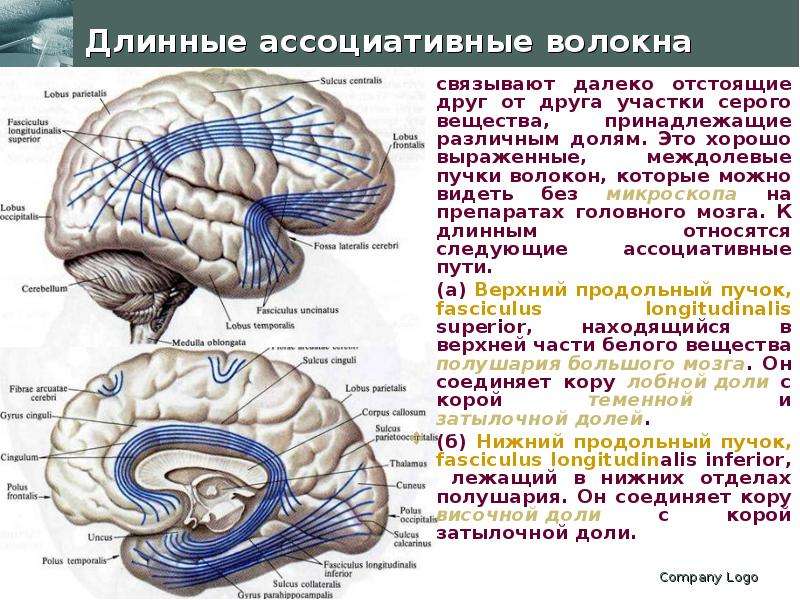
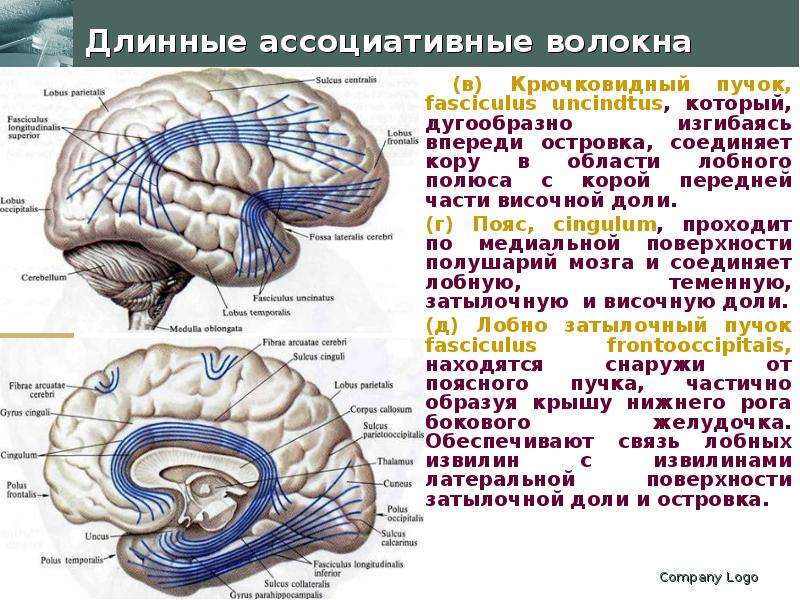
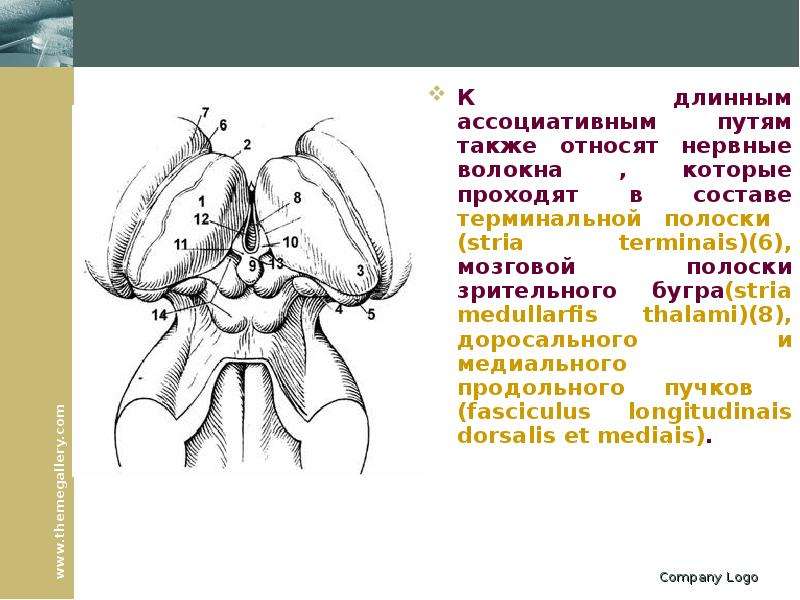
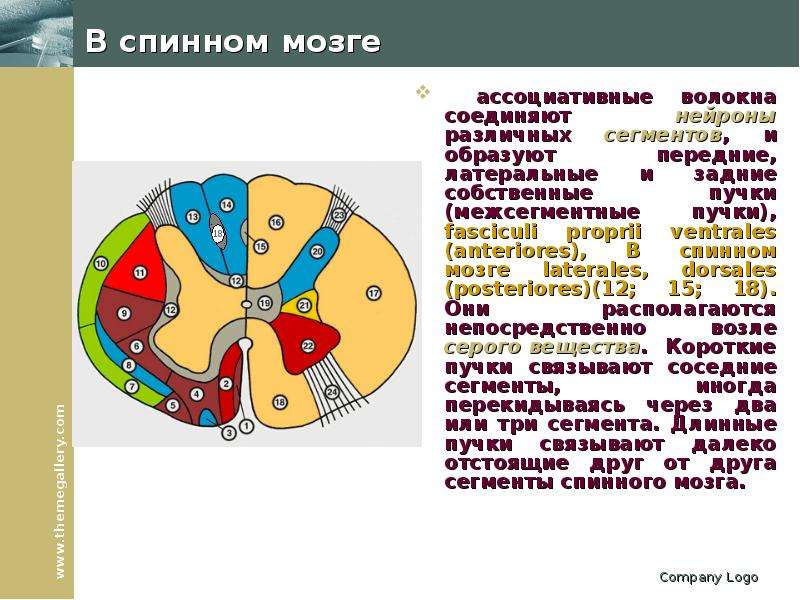
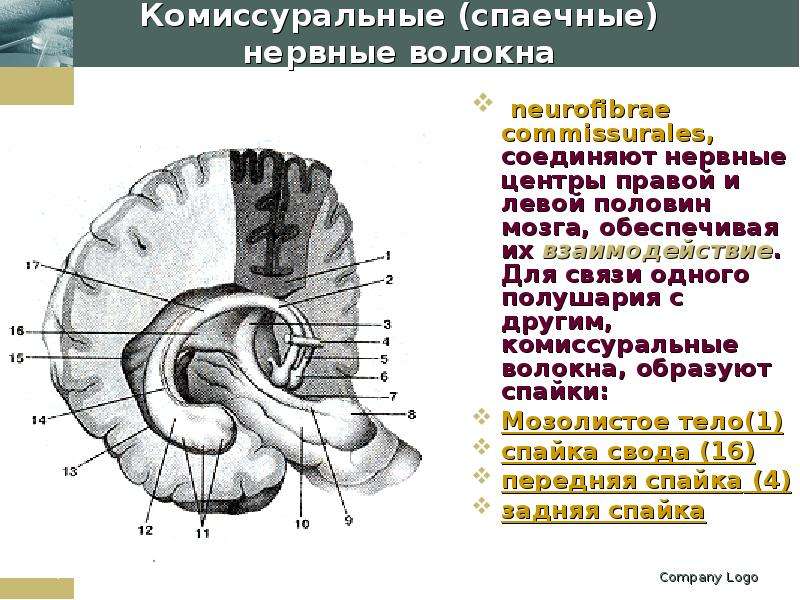
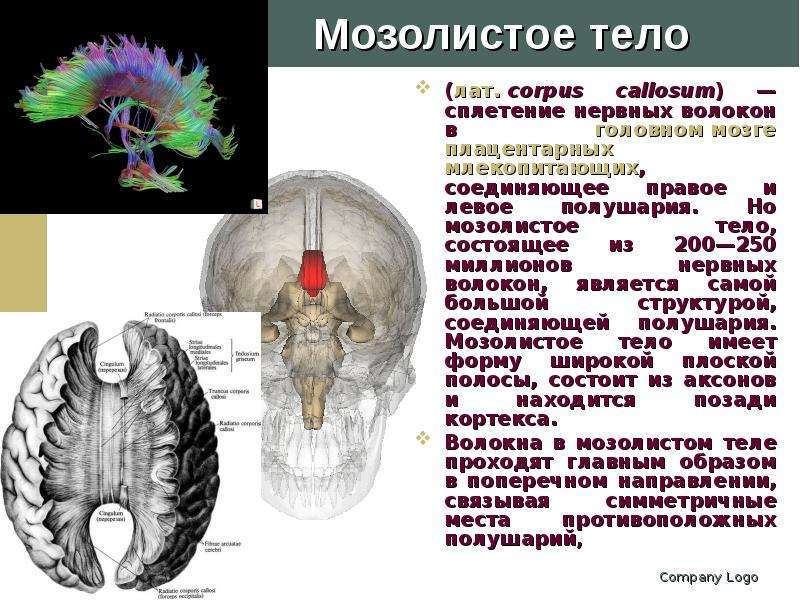
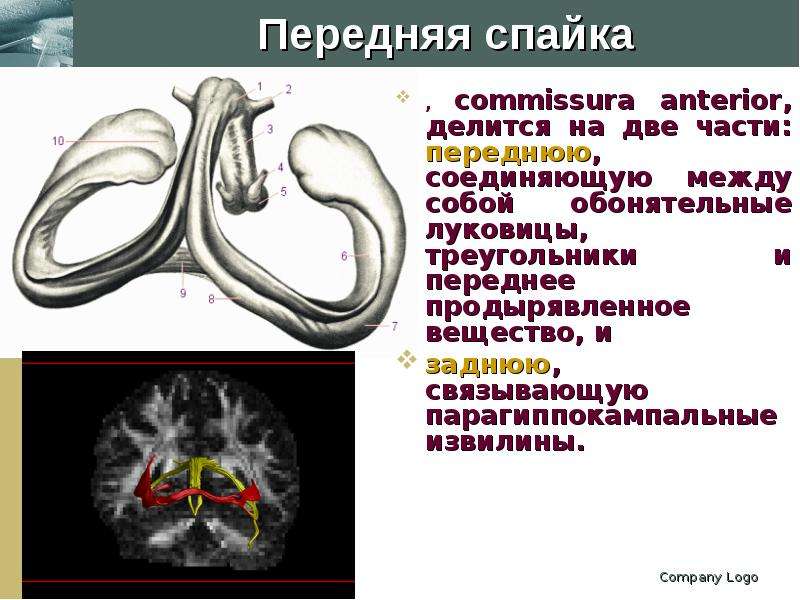
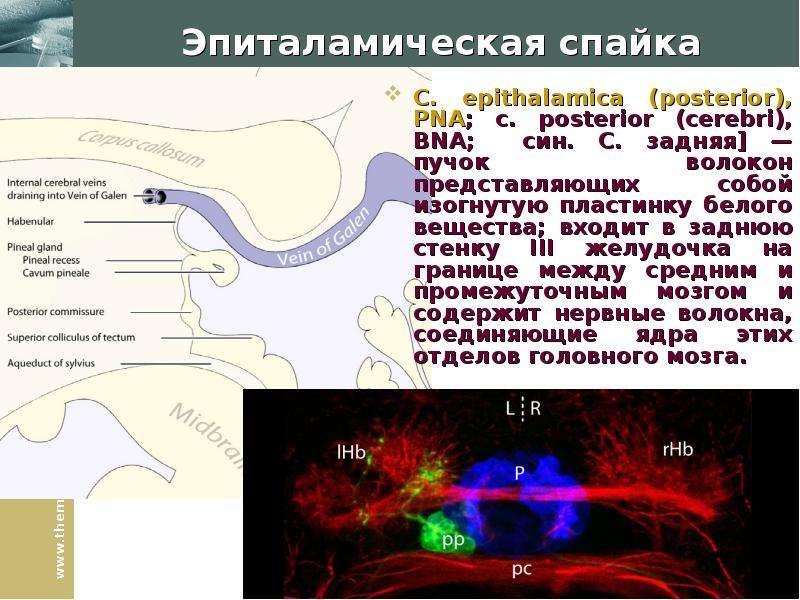
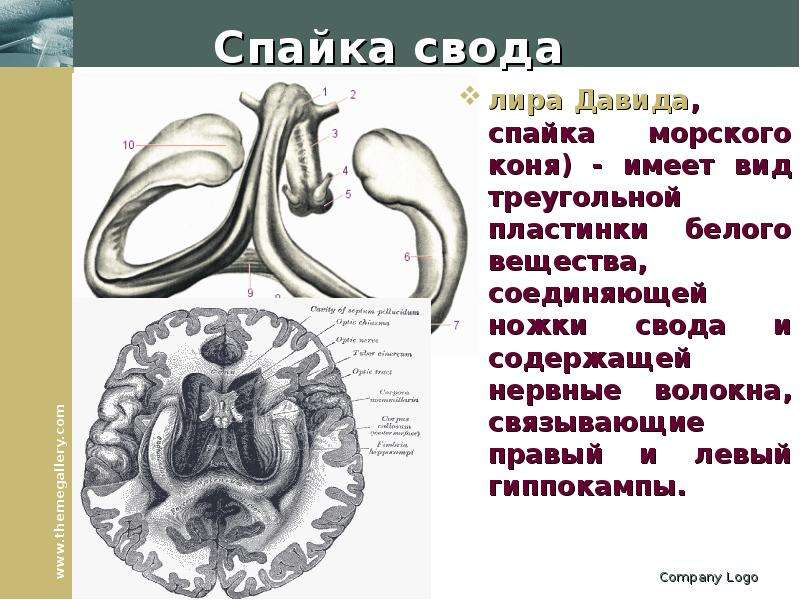
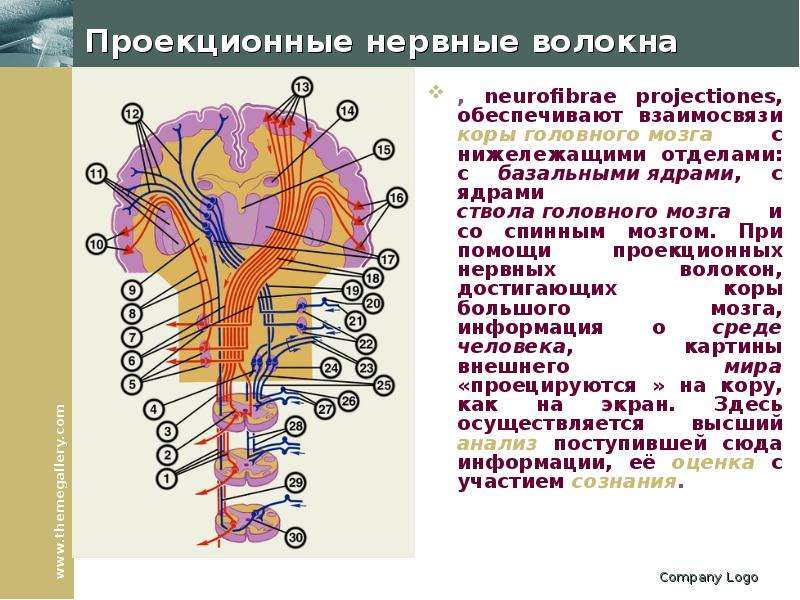
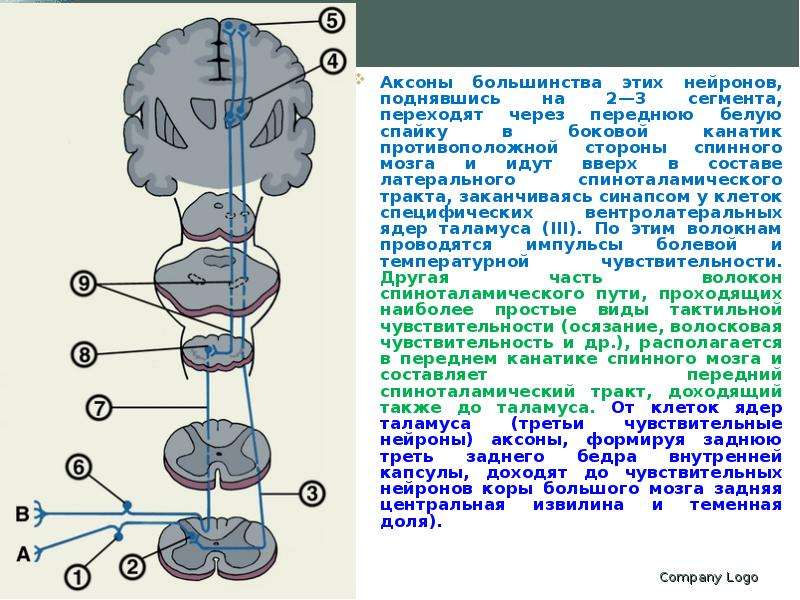
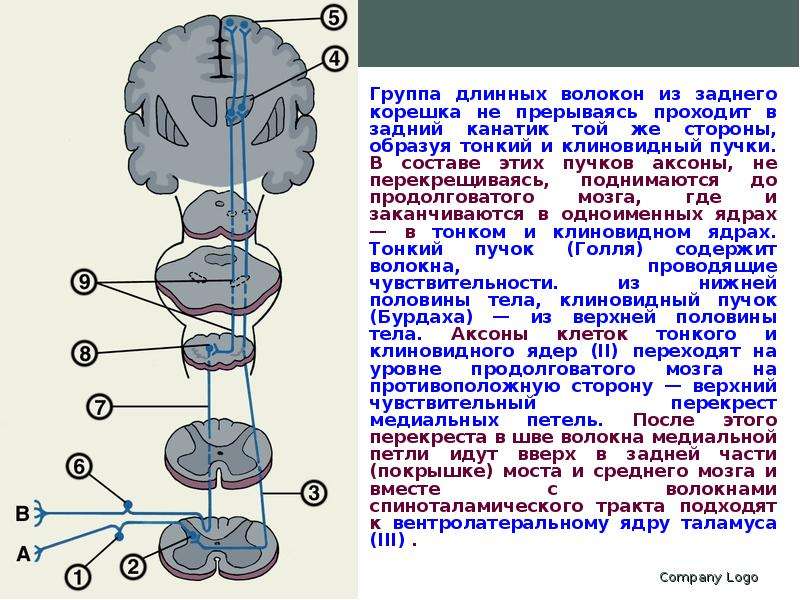
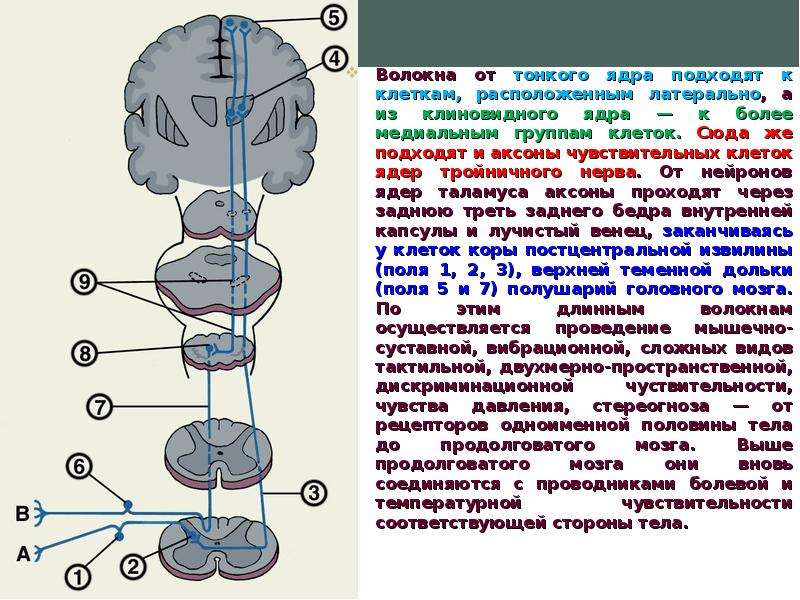
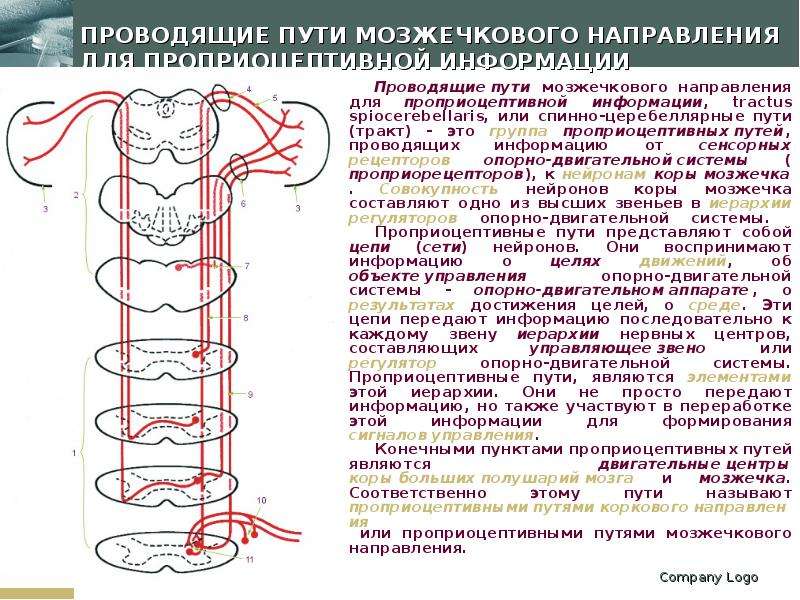
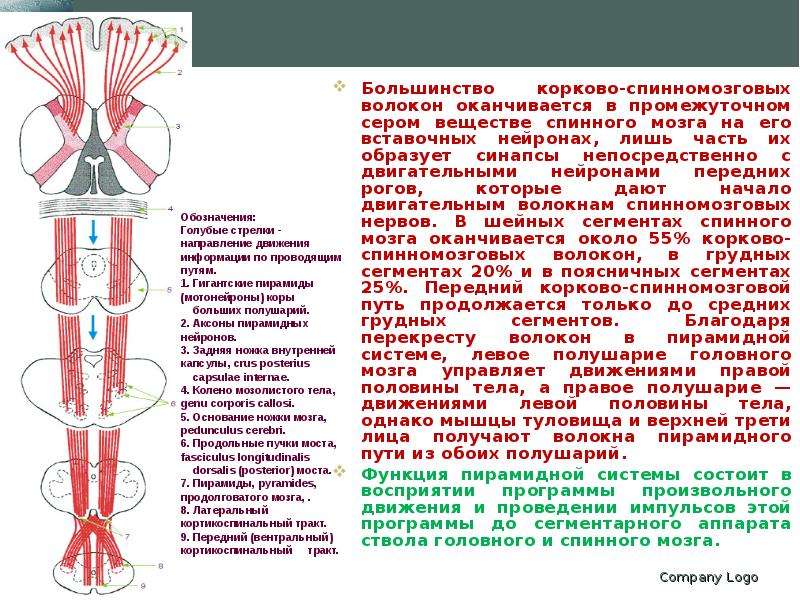
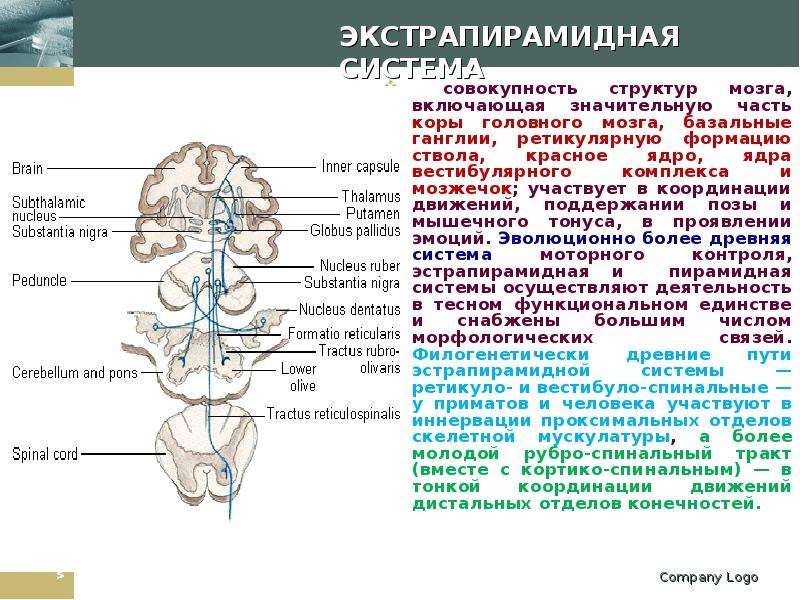
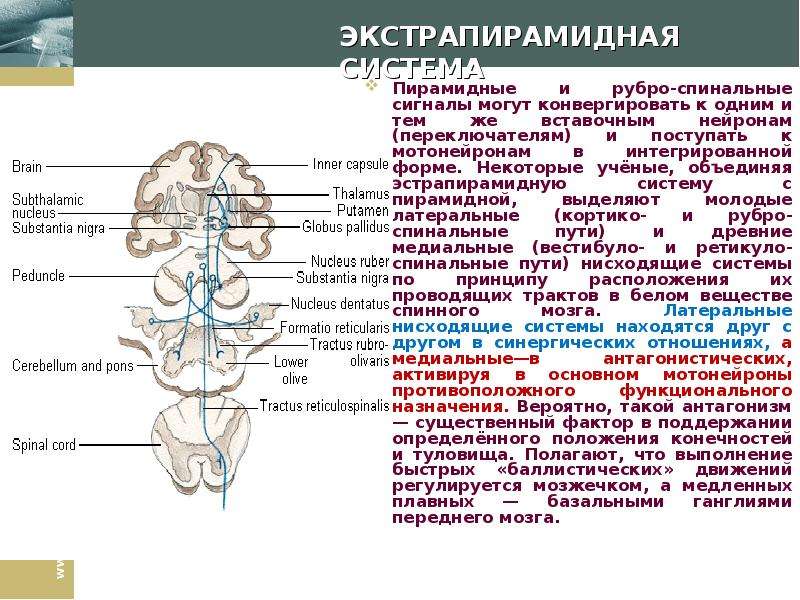
Описание слайда:
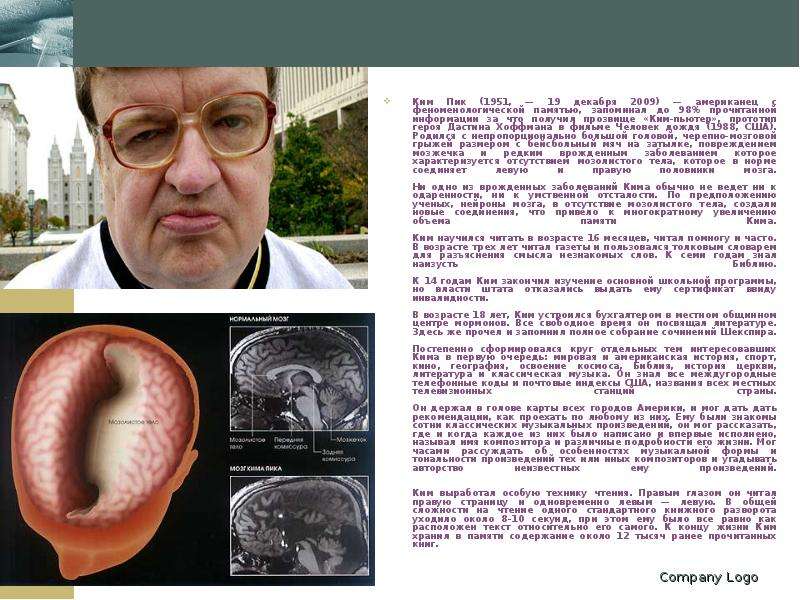
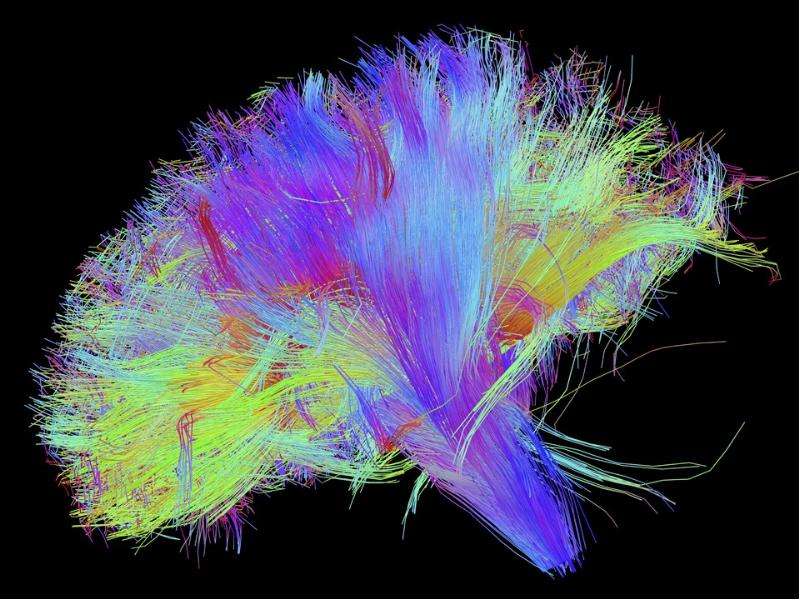
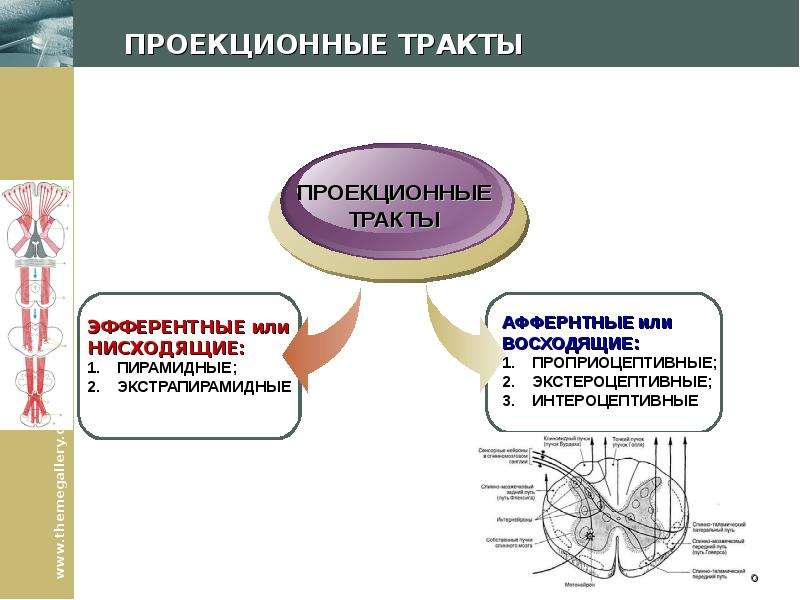
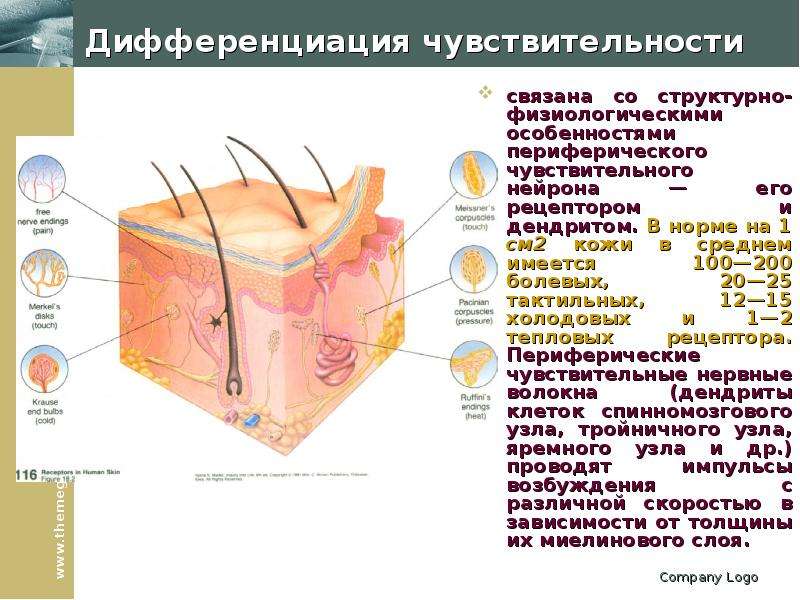
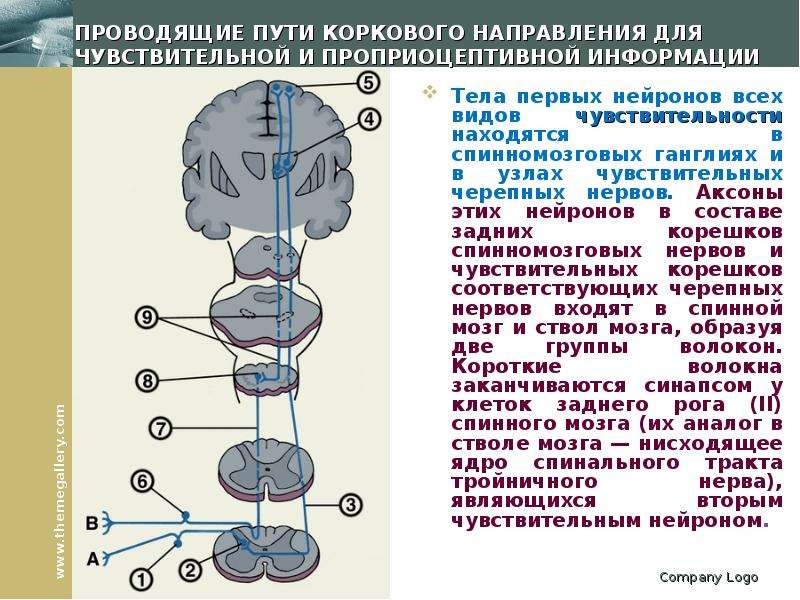
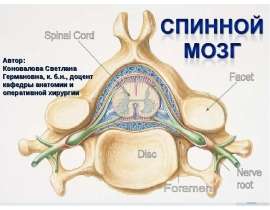
Ким Пик (1951, — 19 декабря 2009) — американец с феноменологической памятью, запоминал до 98% прочитанной информации за что получил прозвище «Ким-пьютер», прототип героя Дастина Хоффмана в фильме Человек дождя (1988, США). Родился с непропорционально большой головой, черепно-мозговой грыжей размером с бейсбольный мяч на затылке, повреждением мозжечка и редким врожденным заболеванием которое характеризуется отсутствием мозолистого тела, которое в норме соединяет левую и правую половинки мозга. Ни одно из врожденных заболеваний Кима обычно не ведет ни к одаренности, ни к умственной отсталости. По предположению ученых, нейроны мозга, в отсутствие мозолистого тела, создали новые соединения, что привело к многократному увеличению объема памяти Кима. Ким научился читать в возрасте 16 месяцев, читал помногу и часто. В возрасте трех лет читал газеты и пользовался толковым словарем для разъяснения смысла незнакомых слов. К семи годам знал наизусть Библию. К 14 годам Ким закончил изучение основной школьной программы, но власти штата отказались выдать ему сертификат ввиду инвалидности. В возрасте 18 лет, Ким устроился бухгалтером в местном общинном центре мормонов. Все свободное время он посвящал литературе. Здесь же прочел и запомнил полное собрание сочинений Шекспира. Постепенно сформировался круг отдельных тем интересовавших Кима в первую очередь: мировая и американская история, спорт, кино, география, освоение космоса, Библия, история церкви, литература и классическая музыка. Он знал все междугородные телефонные коды и почтовые индексы США, названия всех местных телевизионных станций страны. Он держал в голове карты всех городов Америки, и мог дать дать рекомендации, как проехать по любому из них. Ему были знакомы сотни классических музыкальных произведений, он мог рассказать, где и когда каждое из них было написано и впервые исполнено, называл имя композитора и различные подробности его жизни. Мог часами рассуждать об особенностях музыкальной формы и тональности произведений тех или иных композиторов и угадывать авторство неизвестных ему произведений. Ким выработал особую технику чтения. Правым глазом он читал правую страницу и одновременно левым — левую. В общей сложности на чтение одного стандартного книжного разворота уходило около 8-10 секунд, при этом ему было все равно как расположен текст относительно его самого. К концу жизни Ким хранил в памяти содержание около 12 тысяч ранее прочитанных книг. Ким Пик (1951, — 19 декабря 2009) — американец с феноменологической памятью, запоминал до 98% прочитанной информации за что получил прозвище «Ким-пьютер», прототип героя Дастина Хоффмана в фильме Человек дождя (1988, США). Родился с непропорционально большой головой, черепно-мозговой грыжей размером с бейсбольный мяч на затылке, повреждением мозжечка и редким врожденным заболеванием которое характеризуется отсутствием мозолистого тела, которое в норме соединяет левую и правую половинки мозга. Ни одно из врожденных заболеваний Кима обычно не ведет ни к одаренности, ни к умственной отсталости. По предположению ученых, нейроны мозга, в отсутствие мозолистого тела, создали новые соединения, что привело к многократному увеличению объема памяти Кима. Ким научился читать в возрасте 16 месяцев, читал помногу и часто. В возрасте трех лет читал газеты и пользовался толковым словарем для разъяснения смысла незнакомых слов. К семи годам знал наизусть Библию. К 14 годам Ким закончил изучение основной школьной программы, но власти штата отказались выдать ему сертификат ввиду инвалидности. В возрасте 18 лет, Ким устроился бухгалтером в местном общинном центре мормонов. Все свободное время он посвящал литературе. Здесь же прочел и запомнил полное собрание сочинений Шекспира. Постепенно сформировался круг отдельных тем интересовавших Кима в первую очередь: мировая и американская история, спорт, кино, география, освоение космоса, Библия, история церкви, литература и классическая музыка. Он знал все междугородные телефонные коды и почтовые индексы США, названия всех местных телевизионных станций страны. Он держал в голове карты всех городов Америки, и мог дать дать рекомендации, как проехать по любому из них. Ему были знакомы сотни классических музыкальных произведений, он мог рассказать, где и когда каждое из них было написано и впервые исполнено, называл имя композитора и различные подробности его жизни. Мог часами рассуждать об особенностях музыкальной формы и тональности произведений тех или иных композиторов и угадывать авторство неизвестных ему произведений. Ким выработал особую технику чтения. Правым глазом он читал правую страницу и одновременно левым — левую. В общей сложности на чтение одного стандартного книжного разворота уходило около 8-10 секунд, при этом ему было все равно как расположен текст относительно его самого. К концу жизни Ким хранил в памяти содержание около 12 тысяч ранее прочитанных книг.