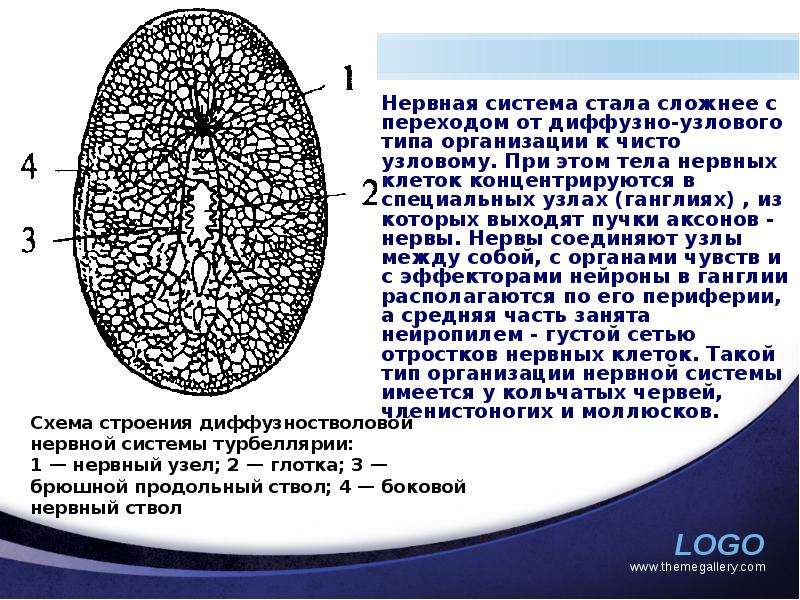
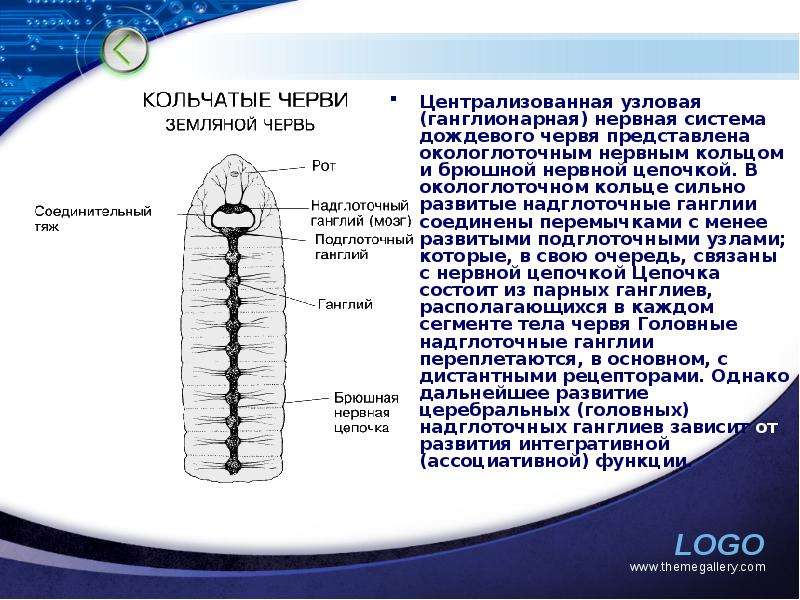
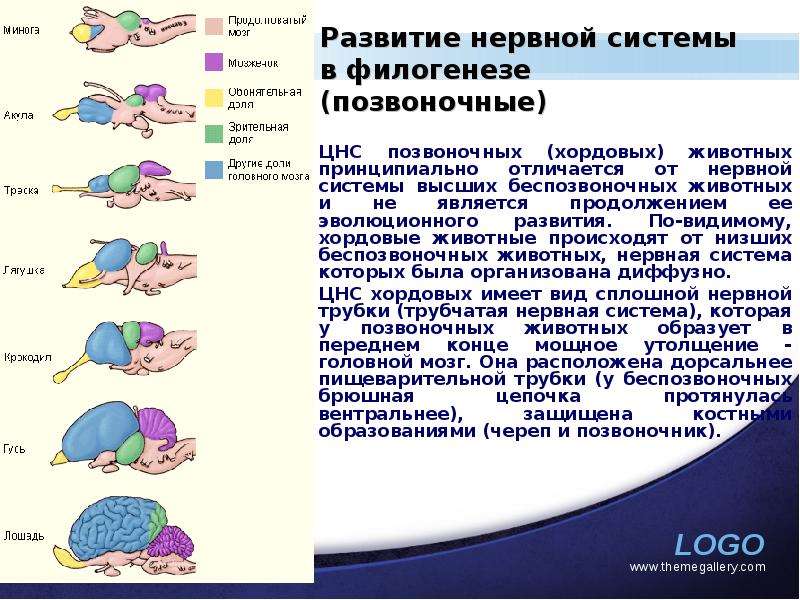
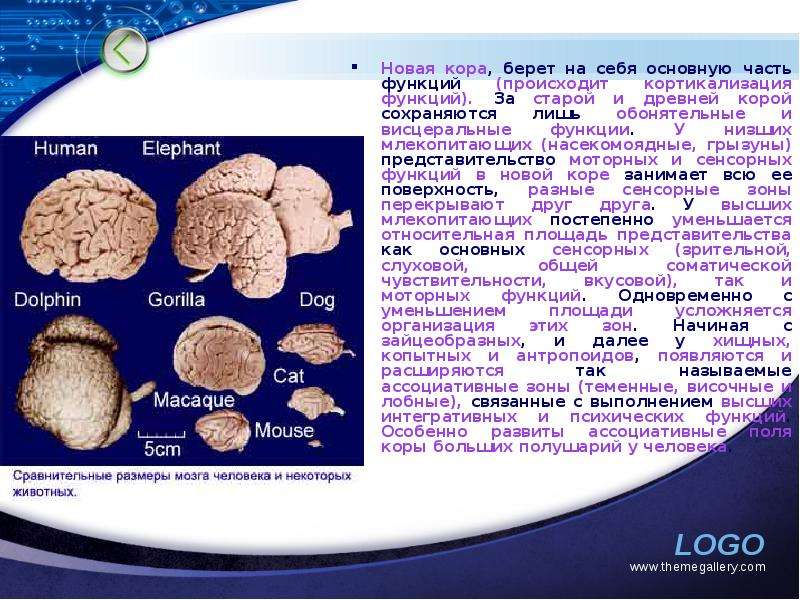
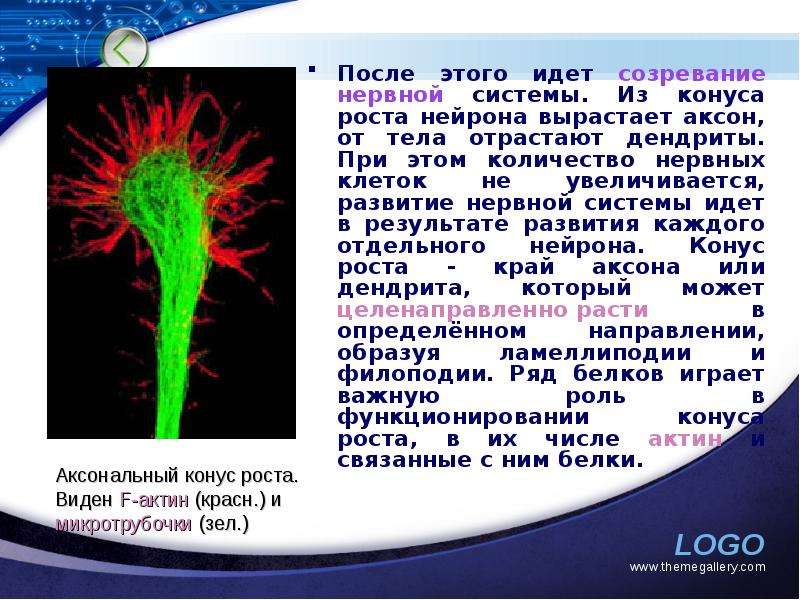
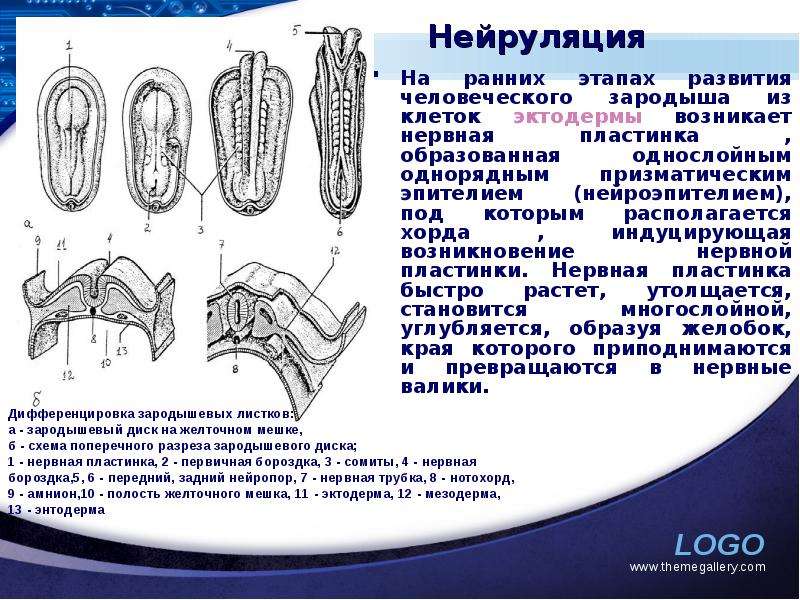
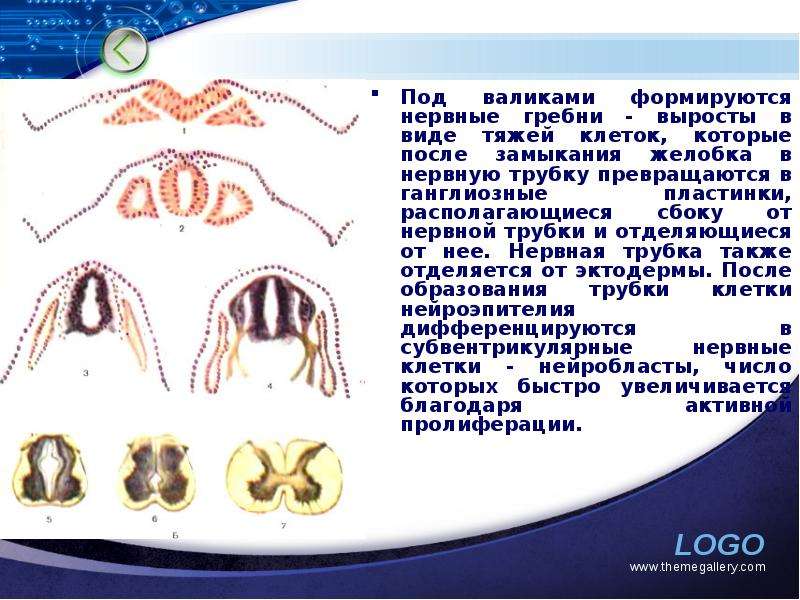
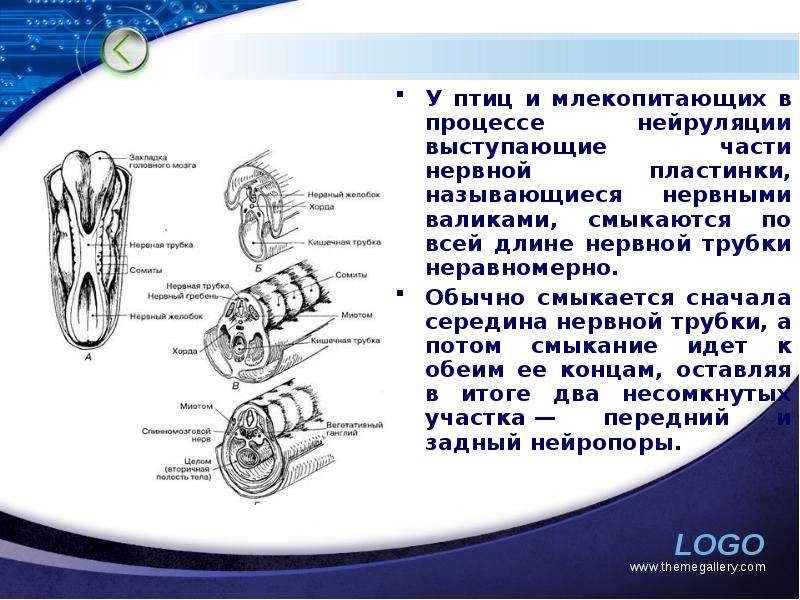
Описание слайда:
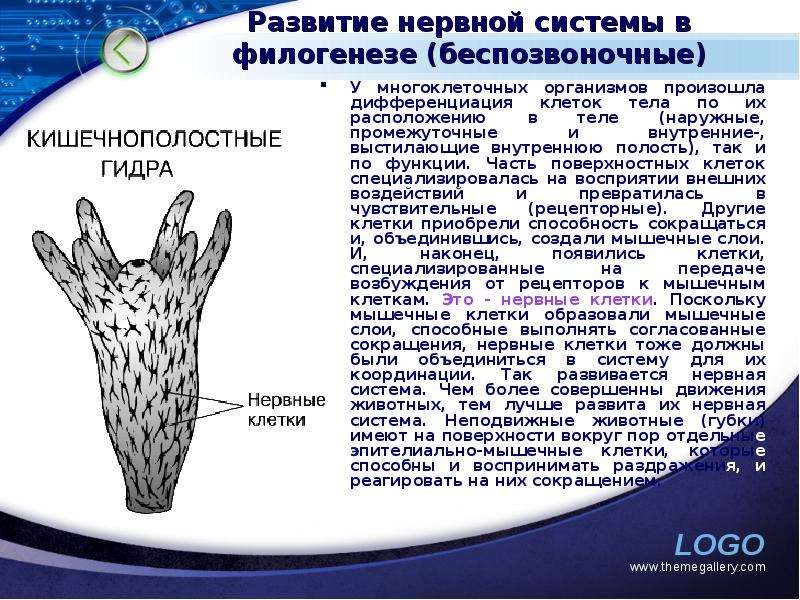
У многоклеточных организмов произошла дифференциация клеток тела по их расположению в теле (наружные, промежуточные и внутренние-, выстилающие внутреннюю полость), так и по функции. Часть поверхностных клеток специализировалась на восприятии внешних воздействий и превратилась в чувствительные (рецепторные). Другие клетки приобрели способность сокращаться и, объединившись, создали мышечные слои. И, наконец, появились клетки, специализированные на передаче возбуждения от рецепторов к мышечным клеткам. Это - нервные клетки. Поскольку мышечные клетки образовали мышечные слои, способные выполнять согласованные сокращения, нервные клетки тоже должны были объединиться в систему для их координации. Так развивается нервная система. Чем более совершенны движения животных, тем лучше развита их нервная система. Неподвижные животные (губки) имеют на поверхности вокруг пор отдельные эпителиально-мышечные клетки, которые способны и воспринимать раздражения, и реагировать на них сокращением. У многоклеточных организмов произошла дифференциация клеток тела по их расположению в теле (наружные, промежуточные и внутренние-, выстилающие внутреннюю полость), так и по функции. Часть поверхностных клеток специализировалась на восприятии внешних воздействий и превратилась в чувствительные (рецепторные). Другие клетки приобрели способность сокращаться и, объединившись, создали мышечные слои. И, наконец, появились клетки, специализированные на передаче возбуждения от рецепторов к мышечным клеткам. Это - нервные клетки. Поскольку мышечные клетки образовали мышечные слои, способные выполнять согласованные сокращения, нервные клетки тоже должны были объединиться в систему для их координации. Так развивается нервная система. Чем более совершенны движения животных, тем лучше развита их нервная система. Неподвижные животные (губки) имеют на поверхности вокруг пор отдельные эпителиально-мышечные клетки, которые способны и воспринимать раздражения, и реагировать на них сокращением.