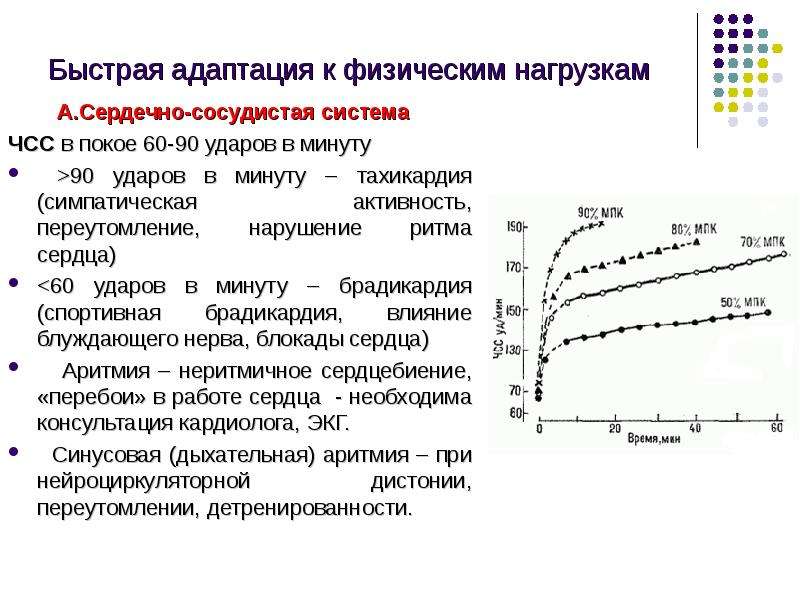
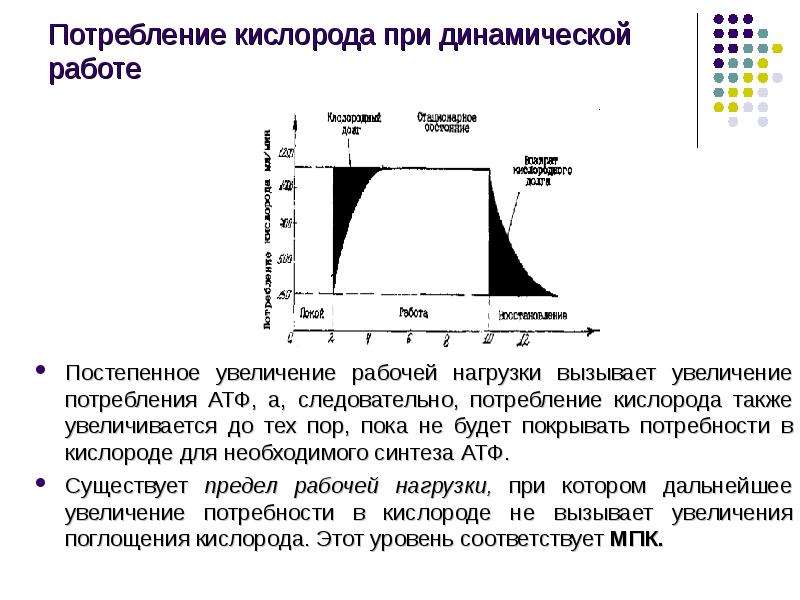
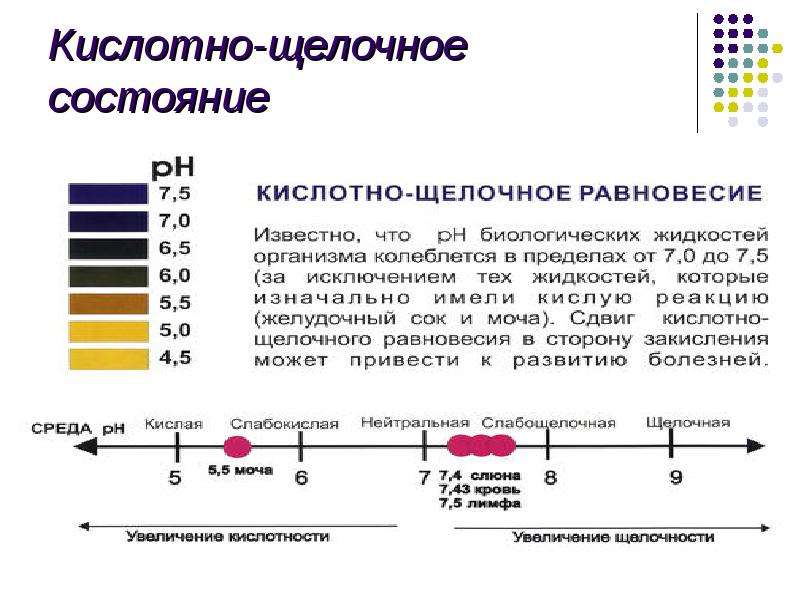
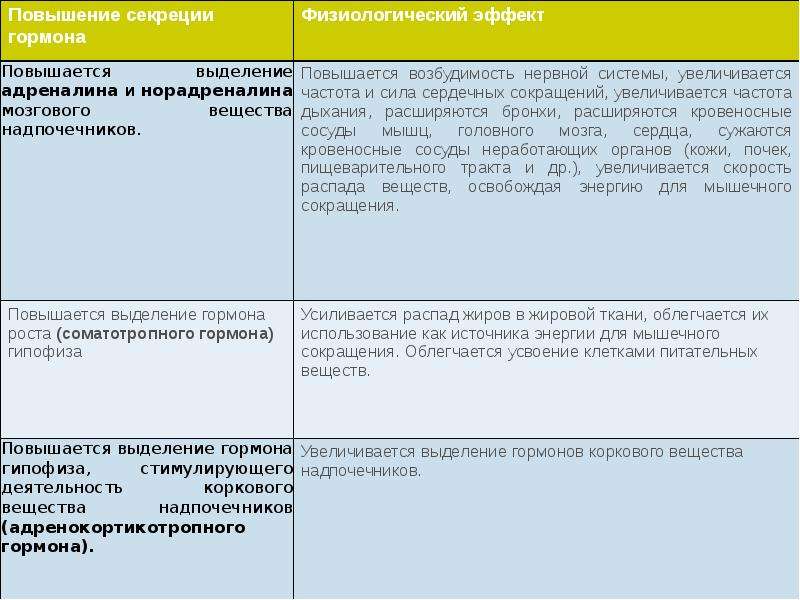
Описание слайда:
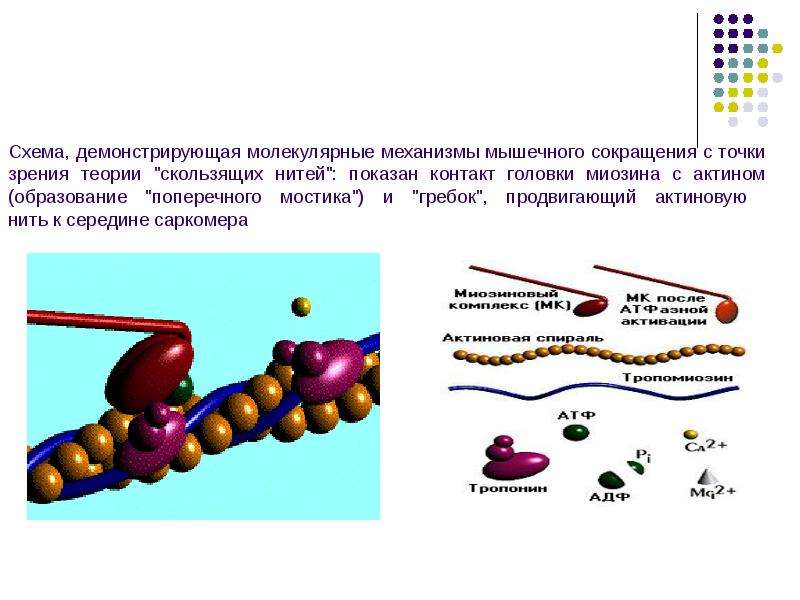
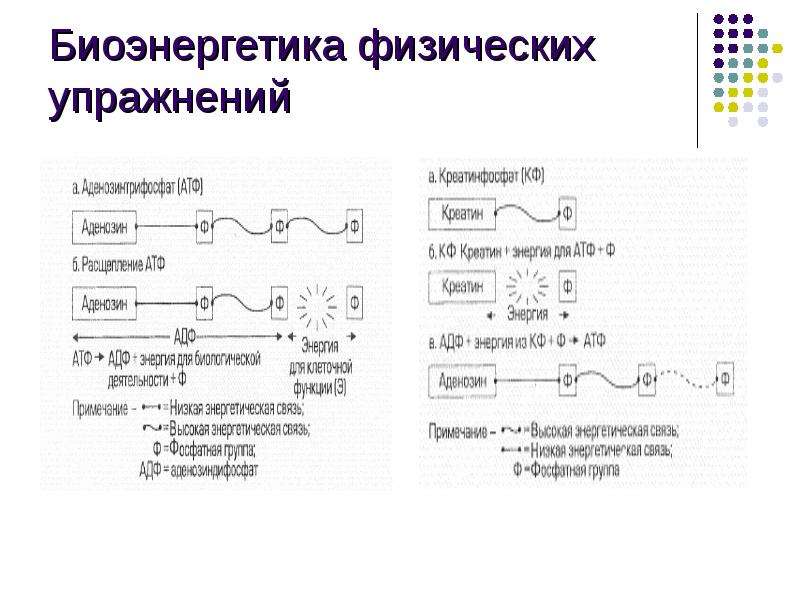
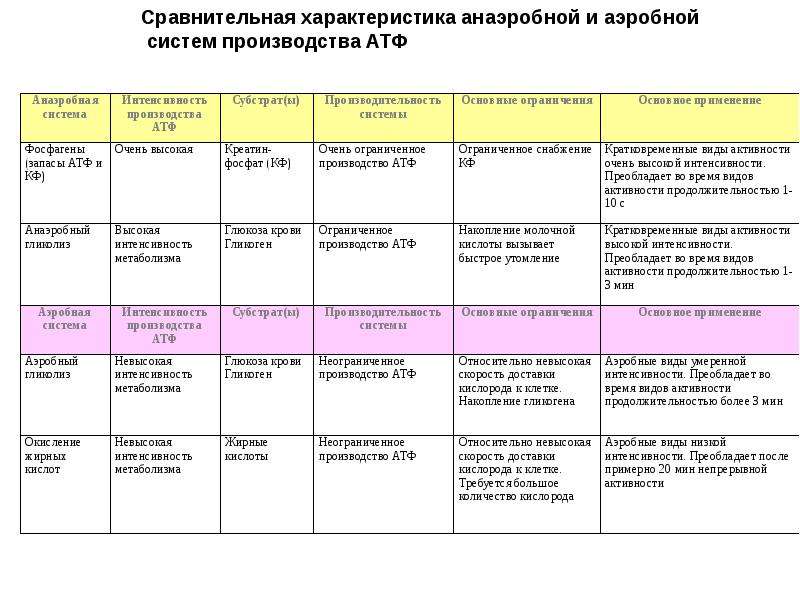
Миозиновые нити несут множество таких «головок» . Как видно из рисунка, в данной реакции кроме актина и миозина принимают участие ещё несколько компонентов. В первую очередь это тропомиозиновые нити, лежащие в желобках, образованных спирально скрученной парой актиновых нитей. Считается, что в состоянии покоя тропомиозин расположен так, что препятствует контакту головки миозина с актином. Для того, чтобы в случае необходимости "открыть доступ", существует так называемый "кальциевый переключатель": через регулярные промежутки актиновые нити несут на себе молекулы тропонина. При связывании с Са(2+) они деформируются таким образом, что смещают тропомиозиновую нить вглубь желобка между актиновыми мономерами, в результате чего контакт миозина с актином становится возможным. Как известно, активация миоцита начинается с деполяризации его мембраны. В результате этого из цистерн эндоплазматического ретикулума в области Z-пластин в саркоплазму начинают выходить ионы Са(2+). Они, в свою очередь, взаимодействуют с тропонином, который смещает тропомиозин вглубь актиновой спирали. Благодаря этому образуется актомиозиновый комплекс, который иногда называют "поперечным мостиком". Донором энергии для перемещения этого мостика (и, соответственно, самого сокращения) является АТФ, которая расщепляется в присутствии ионов Mg(2+) (см. схему). Предполагается, что АТФ расщепляется миозином уже после гребка, давая таки образом энергию для разделения актомиозинового комплекса. В пользу этой точки зрения говорит существование так называемого трупного окоченения (rigor mortis): после смерти организма содержание АТФ в клетках резко снижается, и поперечные мостики долгое время остаются прикреплёнными к актиновой нити - до тех пор, пока не произойдет автолизис. Миозиновые нити несут множество таких «головок» . Как видно из рисунка, в данной реакции кроме актина и миозина принимают участие ещё несколько компонентов. В первую очередь это тропомиозиновые нити, лежащие в желобках, образованных спирально скрученной парой актиновых нитей. Считается, что в состоянии покоя тропомиозин расположен так, что препятствует контакту головки миозина с актином. Для того, чтобы в случае необходимости "открыть доступ", существует так называемый "кальциевый переключатель": через регулярные промежутки актиновые нити несут на себе молекулы тропонина. При связывании с Са(2+) они деформируются таким образом, что смещают тропомиозиновую нить вглубь желобка между актиновыми мономерами, в результате чего контакт миозина с актином становится возможным. Как известно, активация миоцита начинается с деполяризации его мембраны. В результате этого из цистерн эндоплазматического ретикулума в области Z-пластин в саркоплазму начинают выходить ионы Са(2+). Они, в свою очередь, взаимодействуют с тропонином, который смещает тропомиозин вглубь актиновой спирали. Благодаря этому образуется актомиозиновый комплекс, который иногда называют "поперечным мостиком". Донором энергии для перемещения этого мостика (и, соответственно, самого сокращения) является АТФ, которая расщепляется в присутствии ионов Mg(2+) (см. схему). Предполагается, что АТФ расщепляется миозином уже после гребка, давая таки образом энергию для разделения актомиозинового комплекса. В пользу этой точки зрения говорит существование так называемого трупного окоченения (rigor mortis): после смерти организма содержание АТФ в клетках резко снижается, и поперечные мостики долгое время остаются прикреплёнными к актиновой нити - до тех пор, пока не произойдет автолизис.