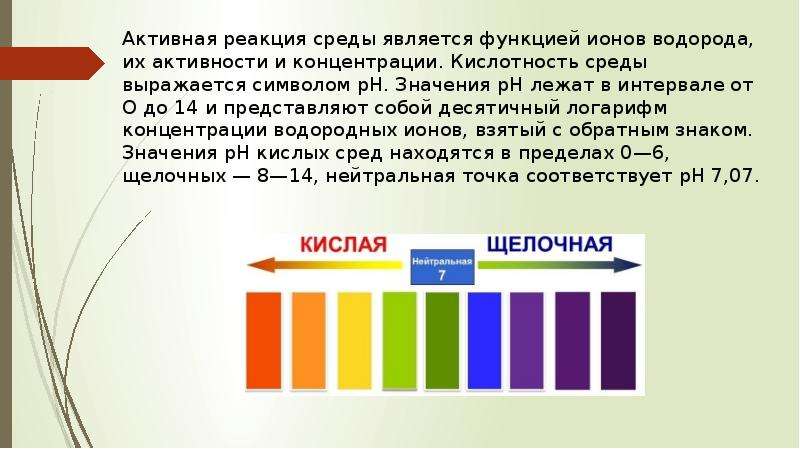
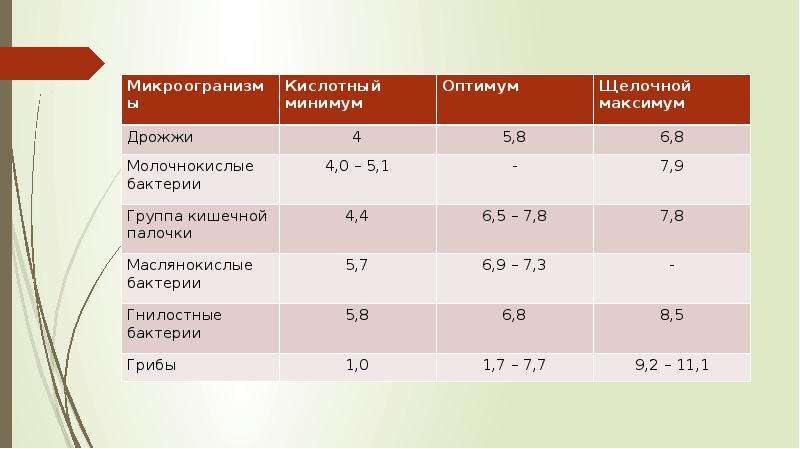
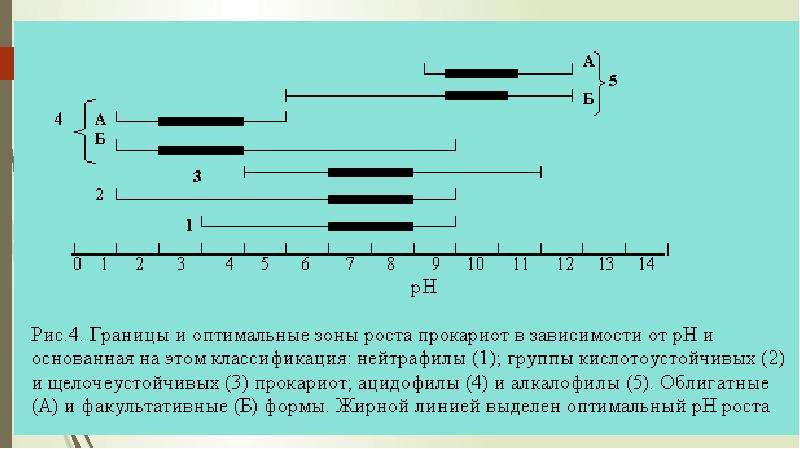
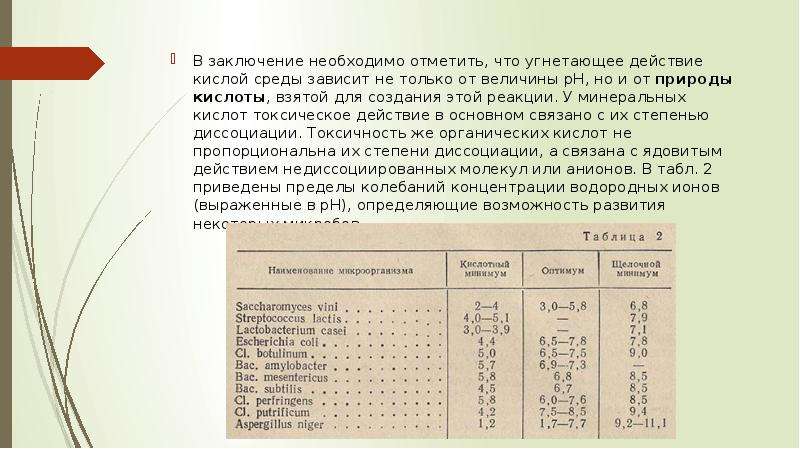
Описание слайда:
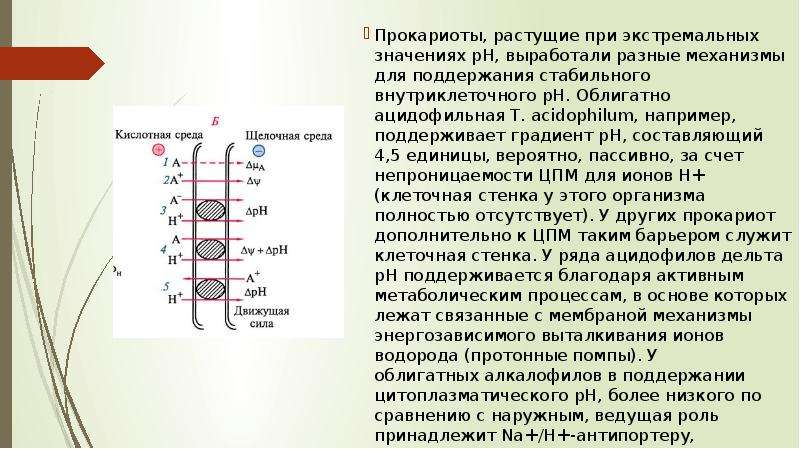
Прокариоты, растущие при экстремальных значениях рН, выработали разные механизмы для поддержания стабильного внутриклеточного рН. Облигатно ацидофильная Т. acidophilum, например, поддерживает градиент рН, составляющий 4,5 единицы, вероятно, пассивно, за счет непроницаемости ЦПМ для ионов Н+ (клеточная стенка у этого организма полностью отсутствует). У других прокариот дополнительно к ЦПМ таким барьером служит клеточная стенка. У ряда ацидофилов дельта рН поддерживается благодаря активным метаболическим процессам, в основе которых лежат связанные с мембраной механизмы энергозависимого выталкивания ионов водорода (протонные помпы). У облигатных алкалофилов в поддержании цитоплазматического рН, более низкого по сравнению с наружным, ведущая роль принадлежит Na+/H+-aнтипортеру, катализирующему движение внутрь клетки протонов в обмен на ионы натрия, в которых эти бактерии нуждаются. Прокариоты, растущие при экстремальных значениях рН, выработали разные механизмы для поддержания стабильного внутриклеточного рН. Облигатно ацидофильная Т. acidophilum, например, поддерживает градиент рН, составляющий 4,5 единицы, вероятно, пассивно, за счет непроницаемости ЦПМ для ионов Н+ (клеточная стенка у этого организма полностью отсутствует). У других прокариот дополнительно к ЦПМ таким барьером служит клеточная стенка. У ряда ацидофилов дельта рН поддерживается благодаря активным метаболическим процессам, в основе которых лежат связанные с мембраной механизмы энергозависимого выталкивания ионов водорода (протонные помпы). У облигатных алкалофилов в поддержании цитоплазматического рН, более низкого по сравнению с наружным, ведущая роль принадлежит Na+/H+-aнтипортеру, катализирующему движение внутрь клетки протонов в обмен на ионы натрия, в которых эти бактерии нуждаются.